INFECÇÃO EXPERIMENTAL DE CAMUNDONGOS (Mus
musculus) E RATOS
(Rattus
norvegicus) COM Strongyloides
venezuelensis
NAKAI 1 E.S. & AMARANTE1 A.F.T.
(1) Departamento de Parasitologia, Instituto de Biociências,
Universidade Estadual Paulista,
Caixa Postal 510, CEP 18618-000,
Botucatu-SP
Experimental Infection in mice (Mus
musculus) and rats (Rattus
norvegicus)
by Strogyloides venezuelensis
The dynamics of the infections by Strongyloides venezuelensis in mice (Mus musculus) and rats (Rattus norvegicus) was studied, as well as the distribution of the parasites in the small intestine of mice. In a first experiment, 12 mice (Group 1) and 12 rats (Group 2) were infected with 500 third stage larvae (L 3 ), while other 12 mice (Group 3) and 12 rats (Group 4) were infected with 4000 L 3 of S. venezuelensis. Fecal samples were collected daily for fecal egg counting (FEC). Seven days after the infection, six animals of each group was sacrificed to determine the worm number. The number of eggs in the uterus and the body length were determined in 10 parasite females recovered from each one of those animals sacrificed. The patent period of the infection was of 79 days in the rats of the group infected with 500 L 3 and of 70 days in the group infected with 4000 L 3 . In the mice the patent periods were of 6 and 9 days in the groups infected with 500 and 4000 L 3 , respectively. The number of eggs in the uterus of the females was similar in the four groups of animals. The medium length of the females was larger in groups 1 and 2, which were infected with the smallest number of larvae. In the second experiment, four groups of mice, with ten animals each, were infected, respectively, with 500, 1000, 2000 and 4000 L 3 . Seven days after the infection the animals were sacrificed and the small intestine was removed and cut in segments of 7 cm. In all groups the largest count of parasites was registered in the first and in the second segment of the small intestine. The last portions presented an insignificant number of nematodes.
KEY WORDS: Strongyloides venezuelensis, Mus musculus, Rattus norvegicus, infection, distribution, dynamics.
INTRODUÇÃO
Os períodos pré-patentes e patentes da infecção por Strongyloides venezuelensis em Mus musculus foram descri-tos em vários artigos (LANTZ et alii, 1998; KOBAYASHI et alii, 1998; KORENAGA et alii, 1995; MARTINS et alii, 2000). No entanto, não foram realizados estudos sobre a dinâmica das infecções por esse parasita em Rattus norvegicus, os hospedeiros originais desse nematódeo. ARAÚJO (1967a), ao examinar 10 Rattus norvegicus silvestres naturalmente infectados com S. venezuelensis, verificou que 90,8% dos parasitas estavam localizados no sexto inicial do intestino delgado. Em camundongos a distribuição deste helminto no in-testino ainda não foi determinada.
Para elucidar alguns aspectos da biologia de S. venezuelensis em Rattus norvegicus e Mus musculus realizou-se este trabalho que teve por objetivos: (1) determinar a dinâmica das infecções primárias e secundárias por S. venezuelensis nos dois hospedeiros; (2) avaliar o desenvolvimento e a prolificidade das fêmeas partenogenéticas de S. venezuelensis em ratos e em camundongos e (3) determinar a distribuição dos nematódeos no intestino delgado de camundongos.
MATERIAL E MÉTODOS
Strongyloides venezuelensis: A cepa de S. venezuelensis utilizada vem sendo mantida em ratos Wistar no Departamento de Parasitologia desde o seu isolamento em Botucatu-SP no início da década de 80. Coproculturas (ROBERTS & O’SULLIVAN, 1950) para produção de larvas infectantes foram realizadas em placa de Petri e as larvas foram separadas das culturas em aparelho de Baermann conforme descrito em PESSÔA & MARTINS (1982).
Animais e infecção artificial: Foram utilizados camundongos (Mus musculus) machos da linhagem Swiss e ratos (Rattus norvegicus) machos da linhagem Wistar, ambos com 25 dias de idade. Os animais foram inoculados com larvas infectantes (L 3 ) de S. venezuelensis, por via subcutânea.
Delineamento experimental: Dois experimentos foram realizados: o primeiro para estudar a dinâmica das infecções em ratos e em camundongos e o segundo para determinar a localização dos nematódeos no intestino delgado de camundongos.
Experimento I - Dinâmica das infecções por Strongyloides venezuelensis em camundongos (Mus musculus) e ratos (Rattus norvegicus): 12 ratos (Grupo 1) e 12 camundongos (Grupo 2) foram infectados com 500 larvas infectantes (L 3 ) de S. venezuelensis e outros 12 ratos (Grupo 3) e 12 camundongos (Grupo 4) foram infectados com 4000 L 3 . Amostras fecais foram colhidas diariamente de todos os grupos de animais até que ovos de S. venezuelensis deixaram de ser observados nos exames de fezes. Para a colheita das amostras fecais, os animais foram distribuídos aleatoriamente em caixas com fundo telado, com capacidade para seis animais, onde permaneceram durante três horas. As fezes eliminadas pelos seis animais foram colhidas e constituíram uma amostra. Em cada uma das amostras foi determinado o número de ovos por grama de fezes (OPG) de acordo com a técnica de GORDON & WHITLOCK (1939), modificada.
Estudo prévio demonstrou que em camundongos o maior número de exemplares de S. venezuelensis é registrado 6-7 dias após a infecção (OLIVEIRA-SEQUEIRA & AMARANTE, 2001). Com isso, sete dias após a inoculação, seis animais de cada um dos grupos foram sacrificados para que fosse determinado o número de nematódeos. Foram colhidas 10 fêmeas partenogenéticas de S. venezuelensis de cada um dos animais sacrificados. O comprimento das fêmeas foi determinado utilizando-se um micrômetro ocular (Zeiss®). Além disso, foi determinado o número de ovos no útero de cada uma das fêmeas.
Os exames de fezes deixaram de ser realizados quando os animais remanescentes (seis em cada grupo) naturalmente eliminaram as infecções, o que foi monitorado pelo exame de fezes. Após 121 dias da primeira infecção, os animais remanescentes dos grupos 1 e 2 foram reinfectados com 500 L 3 e os dos grupos 3 e 4 com 4000 L 3 .
Experimento II - Distribuição de Strongyloides venezuelensis no intestino delgado de camundongos (Mus musculus): Neste experimento foram utilizados quatro grupos de camundongos, cada um com 10 animais. Os animais do grupo 1 foram infectados com 500 L 3 ; os do grupo 2 com 1000 L 3 ; os do grupo 3 com 2000 L 3 e os do grupo 4 com 4000 L 3 . Sete dias após a infecção, os camundongos foram sacrificados.
Obtenção e contagem dos nematódeos do intestino delgado: O intestino delgado foi removido e foi cortado em segmentos de 7 cm e cada um dos segmentos foi colocado, individualmente, em um tubo de ensaio pré-identificado para a recuperação dos nematódeos. O intestino foi invertido em suportes de arame fino e colocado dentro de tubo de ensaio contendo solução fisiológica onde permaneceu incubando durante 4 horas a 37 o C. Decorrido esse tempo, os segmentos intestinais foram retirados dos suportes de arame e a suspensão contendo os nematódeos foi mantida por 6 horas sob refrigeração (4 o C). O sedimento obtido foi acondicionado em frascos com formol 5% e os nematódeos foram contados em placas de Petri sob microscópio estereoscópio.
Análise estatística: Os dados obtidos nos experimentos foram submetidos à análise de variância e as médias foram comparadas pelo Teste de Tukey (OTT, 1992). Os dados referentes às contagens de OPG e de nematódeos foram submetidos à análise de variância após transformação logarítmica (log (x + 1,5)). As associações entre os parâmetros obtidos nos diferentes experimentos foram determinadas calculando-se os coeficientes de correlação de Pearson. No Experimento I foi determinada a correlação de Pearson entre a contagem de vermes, o comprimento das fêmeas e o número de ovos observados no útero. No caso destes dois últimos parâmetros, utilizaram-se os valores médios dos parasitas obtidos de cada animal.
RESULTADOS
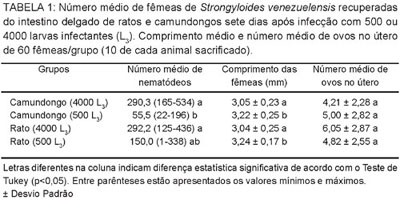
Dinâmica da infecção por Strongyloides venezuelensis em ratos (Rattus norvegicus) e camundongos (Mus musculus): Nos ratos infectados com 500 L 3 e com 4000 L 3 os períodos patentes da infecção foram de 79 dias e de 70 dias, respectivamente (Fig. 1). Nos camundongos infectados com 500 e 4000 L 3 os períodos patentes foram de 6 e 9 dias, respectivamente. Ovos de S. venezuelensis começaram a ser detectados nos exames de fezes dos dois grupos de ratos e no grupo de camundongos infectado com 4000 L 3 cinco dias após a infecção. No grupo de camundongos infectado com 500 L 3 , os ovos foram registrados pela primeira vez nas amostras obtidas seis dias após a infecção. Nos ratos infectados com 4000 L 3 e com 500 L 3 , as maiores contagens de OPG foram registradas oito (96900 OPG) e 11 dias (17700 OPG) após a infecção, respectivamente. Nos camundongos, as contagens mais elevadas foram registradas no sexto (3700 OPG) e no sétimo dia após a infecção (46000 OPG) nos grupos infectados com 500 L 3 e 4000 L 3 , respectivamente.
Seis animais de cada grupo foram sacrificados sete dias após a infecção para que fosse determinada a contagem média de nematódeos no intestino delgado (Tabela 1). As contagens mais elevadas foram registradas nos grupos de animais infectados com 4000 L 3.
Os ratos e os camundongos com cargas parasitárias mais elevadas, infectados com 4000 L 3 , apresentaram fêmeas partenogenéticas com comprimento significativamente inferior ao dos grupos infectados com 500 L 3 (Tabela 1). Já a contagem média de ovos no útero das fêmeas partenogenéticas foi similar nos quatro grupos de animais (Tabela 1). Os coeficientes de correlação entre o número de nematódeos e o comprimento médio das fêmeas foi de – 0,40; entre o número de nematódeos e o número médio de ovos no útero das fêmeas foi de –0,04; e entre o comprimento médio das fêmeas e o número médio de ovos no útero das fêmeas foi de 0,03.
Os seis animais restantes de cada grupo foram reinfectados 121 dias após a infecção primária. Sete dias após a segunda infecção com 500 L 3 , detectou-se 300 OPG nos ratos (Grupo 1) e camundongos (Grupo 2). Nesse mesmo dia, os ratos (Grupo 3) e os camundongos (Grupo 4) reinfectados com 4000 L 3 apresentaram 200 OPG e 100 OPG, respectivamente. No grupo 1 (ratos reinfectados com 500 L 3 ) ainda foram detectados ovos nas fezes por mais cinco dias e no grupo 3 (ratos reinfectado com 4000 L 3 ) foram detectados ovos por mais um dia.
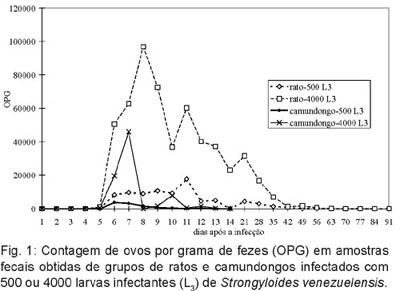
Localização de Strongyloides venezuelensis no intestino delgado de camundongos (Mus musculus): De 65,5% a 72,3% das fêmeas partenogenéticas de S. venezuelensis estavam localizadas nos primeiros 7 cm do intestino delgado nos quatro grupos de camundongos (Figura 2). Um número relativamente elevado, de 23,4% a 31,9%, foi ainda recuperado da segunda porção (7 – 14 cm). Nos dois segmentos iniciais do intestino (0 a 14 cm) foram recuperados de 95,5% a 97,4% do número total de fêmeas partenogenéticas verificado nos quatro grupos de camundongos.
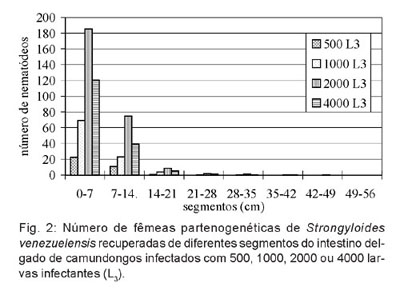
A maior contagem de nematódeos não foi registrada no grupo de camundongos infectado com 2000 L3 (Tabela 2). No entanto, o único grupo que diferiu estatisticamente dos demais foi o infectado com 500 L3, o qual apresentou a contagem mais baixa.
DISCUSSÃO
O período patente da infecção por S. venezuelensis foi bastante distinto nos ratos e nos camundongos tendo sido bem mais longo no primeiro hospedeiro. TAMURA (1993), ao infectar ratos e camundongos com 10000 L3 de S. venezuelensis, verificou que o período patente da infecção foi de 14 dias em camundongos e de 24 dias em ratos e que as contagens máximas de OPG nesses hospedeiros foram, respectivamente, de 2000 e 60000 ovos. No presente estudo o período patente em ratos (de 70 a 79 dias) foi muito superior ao observado por TAMURA (1993). Por outro lado, o período patente em camundongos foi inferior (de 6 a 9 dias). As contagens máximas de OPG registradas neste trabalho em ambos hopedeiros foram mais elevadas do que as obtidas por TAMURA (1993).
De acordo com GROVE & NORTHERN (1989) os ratos e não os camundongos são os hospedeiros naturais de Strongyloides ratti. O mesmo provavelmente acontece com S. venezuelensis. Em estudo realizado na Hungria, KRISKA (1993) encontrou seis espécies de nematódeos e cinco espécies de cestódeos parasitando uma amostra de 590 camundongos (Mus musculus) capturados em residências. Em nenhum dos animais, KRISKA (1993) encontrou parasitas do gênero Strongyloides. Por outro lado, em ratos de esgoto (Rattus norvegicus), ARAÚJO (1967b) verificou que 50% dos animais de uma amostra de 160 ratos, procedentes da cidade de São Paulo, estavam parasitados por S. venezuelensis. Portanto, é provável que na natureza S. venezuelensis seja encontrado parasitando apenas ratos. Isto explicaria as contagens mais elevadas de OPG e os períodos patentes da infecção mais longos nos ratos indicando uma melhor adaptação de S. venezuelensis a estes hospedeiros.
Ao infectar ratos e camundongos com 10000 L3, TAMURA (1993) verificou que em ratos as contagens máximas de OPG foram proporcionais às doses infectantes tendo sido de 7040, 13320, 38400 e 58400 ovos em ratos infectados com 200, 1000, 5000 e 10000 larvas, respectivamente. Por outro lado, o período patente diminuiu a medida que o inóculo aumentou tendo sido de 59, 35, 28 e 28 dias nos ratos infectados com 200, 1000, 5000 e 10000 larvas, respectivamente. No presente estudo, as contagens de OPG também foram proporcionais aos inóculos utilizados, porém os períodos patentes apresentaram durações similares.
Em estudo sobre S. ratti em ratos, KIMURA et alii (1999) observaram dois picos na eliminação de ovos; o maior, com a máxima eliminação de ovos, 7-8 dias após a infecção e um menor por volta de 25 dias após a infecção. No presente trabalho o ápice na eliminação de ovos também ocorreu por volta do oitavo dia após a infecção no grupo de ratos infectado com 4000 L3. Nesse grupo, outras duas elevações nas contagens de OPG foram observadas: no 11o e no 21o dia após a infecção. Já o grupo de ratos infectado com 500 L3 apresentou o ápice nas contagens 11 dias após a infecção. De acordo com KIMURA et alii (1999), a Segunda elevação na contagem de OPG foi produzido por vermes adultos localizados no ceco e no colo. Os autores sugerem que os vermes uma vez eliminados do intestino delgado se estabelecem no intestino grosso. Outros estudos terão que ser realizados para verificar se esse mesmo fenômeno também ocorre com S. venezuelensis em camundongos.
Os períodos pré-patente e patente da infecção por S. venezuelensis verificados em camundongos neste trabalho foram similares aos observados em camundongos das linhagens BALB/c (LANTZ et alii, 1998), C57BL/6 (LANTZ et alii, 1998; KOBAYASHI et alii, 1998; KORENAGA et alii, 1995) e NIH (MARTINS et alii, 2000).
Infecções secundárias por S. venezuelensis não produziram infecções patentes em camundongos BALB/c inoculados com 500 L3 quando o desafio foi realizado 14 dias após a infecção primária (SATO & TOMA, 1990). Já no presente trabalho tanto os ratos quanto os camundongos apresentaram infecção patente ao serem inoculados 121 dias após a infecção primária. Porém, as contagens de OPG foram muito inferiores às observadas na infecção primária. Resultado similar foi obtido por DAWKINS et alii (1980) ao reinfectarem um grupo de camundongos C57BL/6 com 3000 L3 de S. ratti quatro semanas após uma infecção prévia com 400 L3. Nesse caso, o número de vermes recuperados representou menos de um quinto do número recuperado na infecção primária. Entretanto, em um outro experimento, realizado pelos mesmos autores (DAWKINS et alii., 1980), não ocorreram infecções patentes em três linhagens de camundongos previamente infectados com 400 L3 de S. ratti e submetidos a desafio cinco semanas depois com o mesmo número de larvas (400 L3). Aparentemente, para que ocorra o estabelecimento de uma infecção secundária, é necessária uma dose mais elevada de larvas no desafio ou um intervalo de tempo grande entre a infecção primária e a secundária, a exemplo do que ocorreu no Experimento I.
O comprimento das fêmeas partenogenéticas foi inversamente proporcional à carga parasitária dos animais (r = -0,40), não tendo sido influenciada pela espécie do hospedeiro. Resultados diferentes foram observados por MARTINS et alii (2000) ao infectarem camundongos da linhagem NIH convencionais e "germ-free" com 500 L3 de S. venezuelensis. Os autores verificaram que os "germ free" apresentaram cargas parasitárias mais elevadas e também fêmeas partenogenéticas mais compridas que os camundongos "convencionais" (1,79 mm e 2,05 mm, respectivamente).
O número de ovos no útero das fêmeas não teve relação com a carga parasitária, com a espécie do hospedeiro ou com o comprimento das fêmeas o que pode ser demonstrado pelos baixos coeficientes de correlação. Vale ressaltar que algumas fêmeas não apresentavam ovos no útero, é possível que ainda não tivessem iniciado a sua produção.
Em relação à distribuição de S. venezuelensis verificou-se que aproximadamente 70% dos nematódeos localizaram-se nos 7 cm iniciais do intestino delgado dos camundongos. Resultados similares foram obtidos por ARAÚJO (1967a) em ratos silvestres. O autor verificou que 90,8% dos parasitas estavam localizados no sexto inicial do intestino delgado. Em estudos que envolvam a contagem de S. venezuelensis no intestino delgado, sugere-se o processamento apenas do terço inicial do intestino.
Dos quatro inóculos do parasita utilizados na infecção de camundongos, o que resultou no maior número de parasitas adultos no intestino foi o inóculo de 2000 L3. Quando os animais foram infectados com 4000 L3, a carga parasitária resultante foi inferior do que nas infecções com 2000 L3. É provável que inóculos elevados provoquem uma reação imunológica mais acentuada do hospedeiro resultando na eliminação mais rápida dos parasitas.
AGRADECIMENTOS
Ao CNPq pela bolsa concedida à E. S. Nakai e à Sra. Maria Angela Batista Gomes pela colaboração nas atividades laboratoriais.
Foi avaliada a dinâmica das infecções por Strongyloides venezuelensis em camundongos (Mus musculus) e ratos (Rattus norvegicus) e a distribuição do parasita no intestino delgado de camundongos. Em um primeiro experimento, 12 ratos (Grupo 1) e 12 camundongos (Grupo 2) foram infectados com 500 larvas infectantes (L3) de S. venezuelensis, enquanto outros 12 ratos (Grupo 3) e 12 camundongos (Grupo 4) foram infectados com 4000 L3. Amostras fecais foram colhidas diariamente para a contagem de ovos por grama de fezes (OPG). Seis animais de cada grupo foram sacrificados sete dias após a infecção para determinar a carga parasitária. De cada um desses animais, foram colhidas 10 fêmeas partenogenéticas de S. venezuelensis nas quais determinou-se o número de ovos no útero e o comprimento total do parasita. O período patente da infecção foi de 79 dias nos ratos do grupo infectado com 500 L3 e de 70 dias no grupo infectado com 4000 L3. Nos camundongos os períodos patentes foram de 6 e 9 dias nos grupos infectados com 500 e 4000 L3, respectivamente. O número de ovos no útero das fêmeas foi similar nos quatro grupos de animais. O comprimento médio das fêmeas partenogenéticas foi maior nos grupos 1 e 2, os quais foram infectados com o menor número de larvas (500 L3). No segundo experimento, quatro grupos de camundongos, com dez animais cada, foram infectados, respectivamente, com 500, 1000, 2000 e 4000 L3. Sete dias após a infecção os animais foram sacrificados e o intestino delgado foi removido e cortado em segmentos de 7 cm. Nos 14 cm iniciais do intestino delgado foram recuperados de 95,5% a 97,4% do número total de fêmeas partenogenéticas. As últimas porções apresentaram um número insignificante de nematódeos.
PALAVRAS-CHAVE: Strongyloides venezuelensis, Rattus norvegicus, Mus musculus, infecção, distribuição, dinâmica.
REFERÊNCIAS BIBLIOGRÁFICAS
ARAÚJO, P. (1967a). Distribuição de helmintos no intestino delgado de Rattus novergicus naturalmente infestado. II. Strongyloides ratti Sanground, 1925 e S. venezuelensis Brumpt, 1934. Revista da Faculdade de Farmácia e Bioquímica de São Paulo, 1:179-190.
ARAÚJO, P. (1967b). Helmintos de Rattus novergicus (Berkenhout, 1769) da cidade de São Paulo. Revista da Faculdade de Farmácia e Bioquímica de São Paulo, 5:141-159.
DAWINKS, H.J.S., GROVE, D.I., DUNSMORE, J.D. & MITCHELL, G.F. (1980). Strongyloides ratti: susceptibility to infection and resistence to reinfection in inbred strains of mice as assessed by excretion of larve. International Journal for Parasitology, 10:125-129.
GORDON, H.M. & WHITLOCK, H.V. (1939). A new technique for counting nematode eggs in sheep faeces. Journal of the Council for Scientific and Industrial Research, 12:50-52.
GROVE, D.I. & NORTHERN, C. (1989). Dissociation of the protective immune response in the mouse to Strongyoides ratti. Journal of Helminthology, 63:307-314.
KIMURA, E., SHINTOKU, Y., KADOSAKA, T., FUJIWARA, M.; KONDO, S. & ITOH, M. (1999). A second peak of egg excretion in Strongyloides ratti-infected rats: its origin and biological meaning. Parasitology, 119:221-226.
KOBAYASHI, T., TSUCHIYA, K., HARA, T., NAKAHATA, T., KURUKAWA, M., ISHIWATA, K., UCHIYAMA, F. & NAWA, Y. (1998). Intestinal mast cell response and mucosal defence against Strongyloides venezuelensis in interleukin-3-hyporesponsive mice. Parasite Immunology, 20:279-284.
KORENAGA, M., HITOSHI, Y., TAKATSU, K. & TADA, I. (1995). Cross-resistance between Strongyloides venezue-lensis and S. ratti in mice. Journal of Helminthology, 69:119-123.
KRISKA, T. (1993). Parasitic helminths of house mouse (Mus musculus Linnaeus, 1758) in Hungary. Miscellanea Zoologica Hungarica, 8:13-23.
LANTZ, C.S., BOESIGER, J., SONG, C.H., MACH, N., KOBAYASHI, T., MULLIGAN, R.C., NAWA, Y., DRANOFF, G. & GALLI, S.J. (1998). Role for interleukin-3 in mast-cell and basophil development and in immunity to parasites. Nature, 392:90-93.
MARTINS, W.A., MELO, A.L., NICOLI, J.R., CARA, D.C., CARVALHO, M.A.R., LANA, M.A., VIEIRA, E.C. & FARIAS, L.M. (2000). A method of decontaminating Strongyloides venezuelensis larvae for the study of strongyloidiasis in germ-free and conventional mice. Journal of Medical Microbiology, 49:387-390.
OLIVEIRA-SEQUEIRA, T.C.G. & AMARANTE, A.F.T. (2001). Dynamics of Strongyloides venezuelensis infection and relationship between fecal egg counts and parasite burden in Swiss mice. Revista Brasileira de Medicina Veterinária, 23:99-102.
OTT, R.L. (1992). An introduction to statistical methods and data analysis. 4 ed. Wadsworth Publishing Company, 1051 p.
PESSÔA, S.B. & MARTINS, A.V. (1982). Parasitologia médica. 11 ed. Guanabara Koogan, Rio de Janeiro, 872 p.
ROBERTS, F.H.S. & O´SULLIVAN, S.P. (1950). Methods for egg counts and larval cultures for strongyles infesting the gastrointestinal tract of cattle. Australian Journal of Agricultural Research, 1: 99-102.
SATO, Y. & TOMA, H. (1990). Strongyloides venezuelensis infections in mice. International Journal for Parasitology, 20:57-62.
TAMURA, N. (1993). Studies on availability of Strongyloides venezuelensis for experimental model of strongyloidosis. Bulletin of Nippon Veterinary and Animal Science University, p. 94-96.